L'ALBERO, UN'ENTITA' BIOLOGICARelatore: Prof. Pierre RaimbaultScuola Nazionale d'Ingegneria Francia |
L'ALBERO NEL RINVERDIMENTO DEI TETTIRelatore: Prof. Hans-Joachim LieseckeUniversita' di Hannover Germania |
STABILITA' DEGLI ALBERIRelatore: Prof. Claus MattheckUniversita' di Karlsruhe Germania |
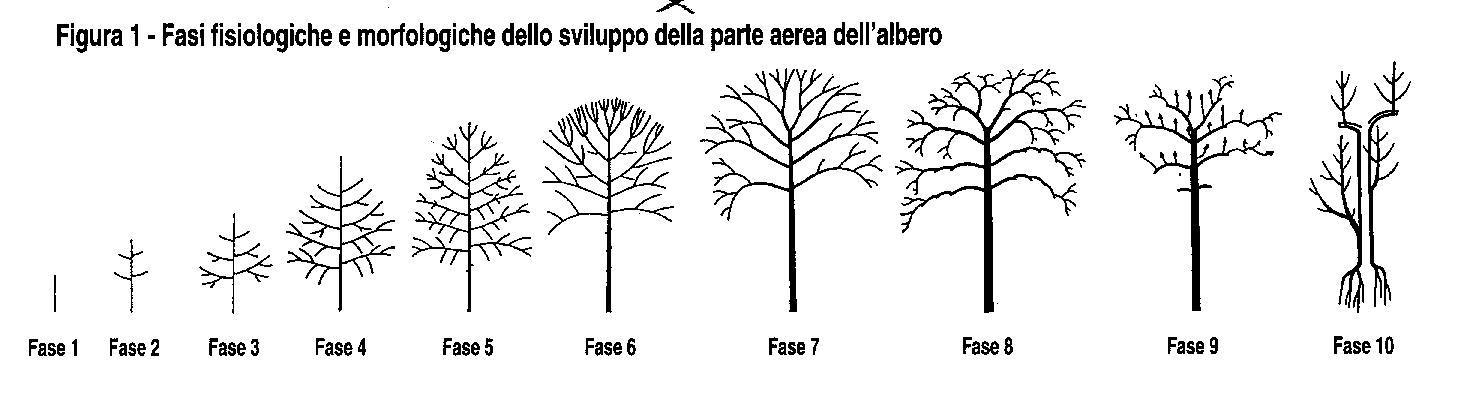
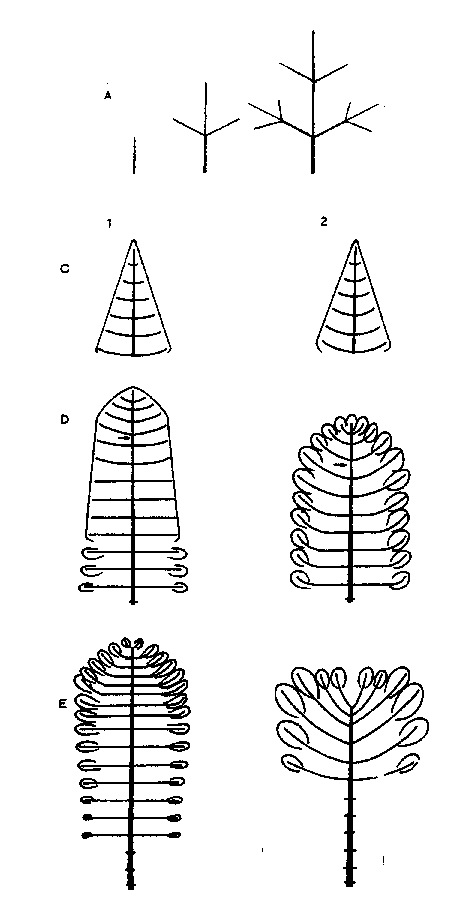 | Le cellule animali vengono sostituite con regolarita', mentre nelle piante le nuove cellule si accumulano esternamente rispetto alle vecchie cellule che restano, vive o morte, nello stesso luogo. I tessuti vengono accumulati dagli alberi in due direzioni: in lunghezza i nuovi germogli vengono aggiunti agli apici dei germogli dell'anno precedente; in spessore gli anelli lignei vivi vengono aggiunti esternamente a quelli vecchi. Questi due tipi di crescita hanno conseguenze rispettivamente sul rinnovo delle branche e sull'evoluzione dell'apparato radicale. Si possono trarre tre conclusioni pratiche. In primo luogo gli animali feriti possono guarire sostituendo le cellule morte con cellule nuove; negli alberi le cellule morte restano definitivamente dove si trovano, a meno che non siano distrutte da funghi, e quelle nuove coprono il legno morto ma non guariscono la ferita in senso stretto: cio' pone il problema della diagnosi patologica meccanica e fisiologica. In secondo luogo, una parte amputata di un animale non puo' essere sostituita, o puo' esserlo solo in casi rari: potare un ramo di un albero provoca una reazione morfogenetica che ripristina la parte tagliata; cio' pone il problema degli effetti della potatura e della risposta fisiologica. In terzo luogo, tutti gli organi di un corpo animale hanno all'incirca la stessa eta'; nelle piante un meristema e' soggetto ad un invecchiamento ontogenetico o a un movimento morfogenetico; indipendentemente dalle condizioni fisiologiche, la morfogenesi muta passando da un esemplare giovane a uno adulto e a uno senescente ma ancora funzionale: cio' solleva la questione dell'invecchiamento fisiologico e ontogenetico nello spazio e nel tempo. |
 | Home page | | Recupero ambientale | | Tecnologie specifiche per la salvaguardia ambientale |